New polio vaccines
Since the 1950s there have been two safe and effective vaccines against poliomyelitis:
- inactivated (killed) polio vaccine – known as IPV or Salk – which is made from wild strains and administered by injection
- live attenuated (weakened) oral polio vaccine – OPV or Sabin – which is given by mouth
OPV has been used in the Global Polio Eradication Initiative to reduce polio cases by more than 99% and to restrict poliovirus circulation to a handful of countries.
But it is a live virus vaccine which can evolve over time into more virulent strains. Because of this many countries – including the UK – have switched to IPV. IPV gives excellent immunity against poliomyelitis but is less effective than OPV in preventing infection and transmission, so both vaccines are needed to successfully eradicate the disease.
Genomic structure of strains for Vero cell vaccines
Poliovirus has a single-stranded RNA genome translated as polyprotein – the four capsid proteins which determine serotype and antigenic structure and are derived from the P1 gene.
Vaccine seeds encode P1 from the three strains used now in IPV as well as the three Sabin strains. The non-structural proteins derive from Sabin 3 and have been modified in the 2A gene in a way that adapts them to Vero cell growth. The non-coding regions which control the infectivity and genetic stability of the strains come from a construct called S19.
There are still several challenges to be overcome to complete and maintain global polio eradication including:
- bio-containment of wild polioviruses – such as those used in IPV production – to help prevent it escaping from facilities into the outside world
- prevention and control of outbreaks caused by OPV strains that have evolved into essentially wild viruses – now and in the future
New vaccines
Because of NIBSC’s long-standing expertise in poliovaccines – and as part of our contribution to global public health – we are involved in three projects to develop new vaccines to help eradicate the disease.

First, we have developed new strains of poliovirus to make IPV without the risk of it accidentally escaping into the human population from a facility. This means it can be made in places where people’s immunity is lower and where sanitation is not ideal – as in many low-cost economies. These strains are now licensed to a major vaccine producer.
A second approach involves engineering the capsid proteins of poliovirus to allow the production of virus-like particles (VLPs) which are antigenically equivalent to live viruses and are stable enough to allow vaccine production using recombinant expression – not using any live virus – which is ideal for the post-eradication world.
Thirdly, we are developing new live attenuated strains which are genetically stable and which are less likely to regain wild-type properties. These would be useful to control outbreaks after vaccination with Sabin strains is stopped.
The strains
The IPV seed strains we have developed can replicate in cell culture – allowing vaccine production – but are effectively non-infectious for humans, and are highly attenuated as a result. Strains are genetically stable so that they should not become infectious again during production. Because of the way they are designed, the strains elicit the same protective immunity as current IPV.
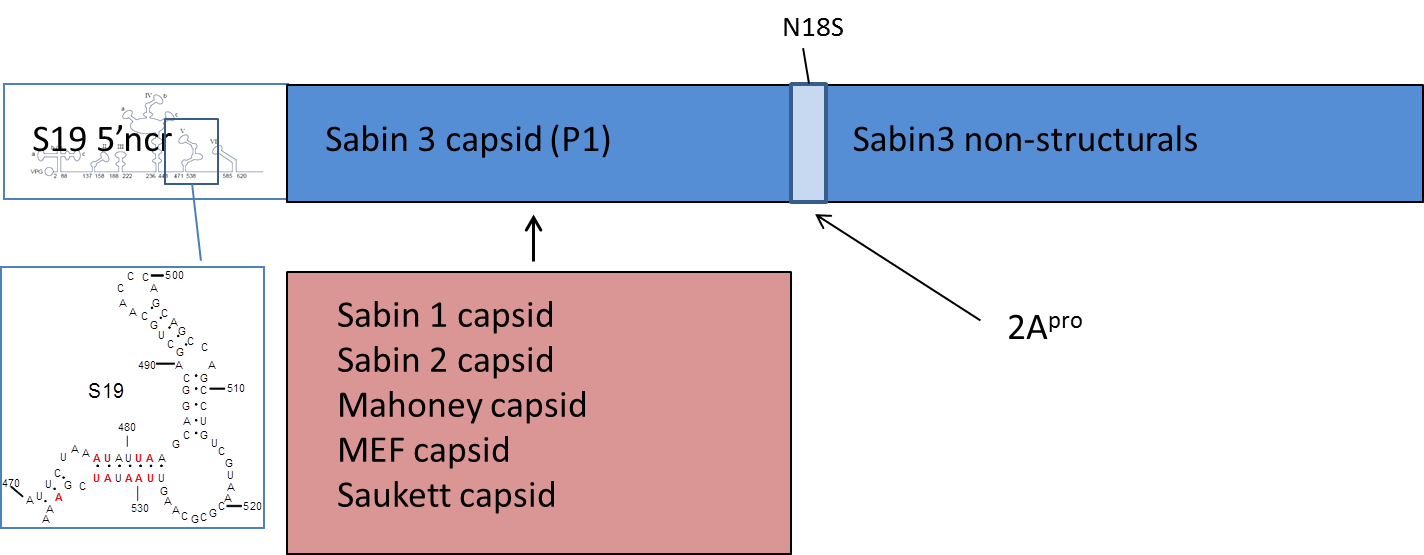
S19-IPV strains
Thermostable poliovirus virus-like particles (VLPs)
VLPs containing no viral RNA – empty capsids – are produced naturally during poliovirus infections. The external surface of a VLP is indistinguishable from that of a virus particle but several internal amino acid chains are disordered. As a result VLPs are immunogenic but conformationally unstable at physiological temperatures.
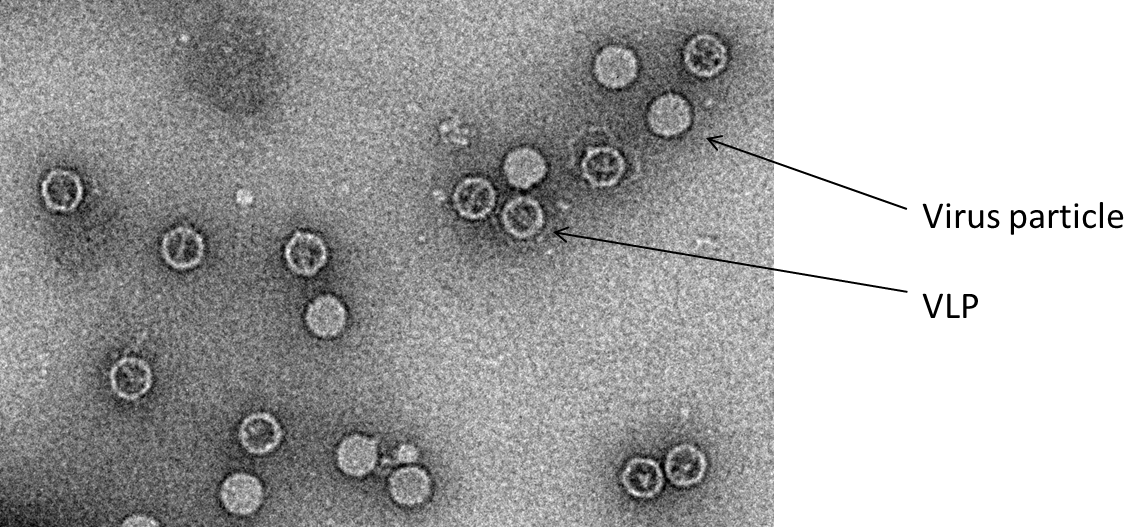
We have identified amino acid substitutions that stabilise VLPs without altering antigenicity and these VLPs appear to be at least as thermostable as the current inactivated – Salk – polio vaccine. We are developing the production of stable VLPs using recombinant mammalian, yeast, bacterial, baculovirus and plant expression systems in collaboration with several UK laboratories:
Genetically-stable OPV strains
The 5’ non-coding region of the poliovirus genome is highly structured. RNA domain V – crucial for both gut infectivity and attenuation – forms part of a larger structure that controls initiation of translation, and the level of attenuation is determined by its thermodynamic stability.
We have modified domain V to make it genetically stable by removing all U-G base-pairs. Any single mutation in base-paired stems will weaken this structure and generate a more attenuated virus.
We have also moved an essential cis-acting replication element (cre) from the P2 region to near the 5’ end to reduce the risk of loss of this part of the genome through recombination. In this way two recombination events are required to replace domain V – a single event would also remove the cre and the resulting genome would be non-infectious.
In a collaboration with the Andino Lab at the University of California, San Francisco, we are also modifying the polymerase to introduce mutations that increase its fidelity and decrease its ability to catalyse homologous recombination.